面向东海生态系统的OSMOSE-ECS模型构建与应用
黄猛,李曰嵩,2,3,4*
(1.上海海洋大学 海洋科学学院,上海 201306;
2.国家远洋渔业工程技术研究中心,上海 201306;
3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海 201306;
4.远洋渔业协同创新中心,上海 201306)
海洋占据了地球表面71%的面积,其中蕴藏了丰富的生物资源,它们不仅能为人类提供优质的蛋白质,同时也是许多工业生产过程中不可或缺的原材料。但是,长久以来的渔业生产活动对海洋渔业资源产生了较大影响[1],过度捕捞和环境污染使得海洋渔业资源发生了严重的衰退,许多渔业发达国家已经注意到了这些问题,并开始寻求海洋渔业发展方式的转变。
在此背景下,基于生态系统的渔业管理方式(ecosystem-based fishery management,EBFM)应运而生[2-3]。许多国际公约和组织机构倡导采用可持续的渔业管理方式实施渔业生产作业,以维持和恢复海洋生态系统的健康状态[4]。为实现这一目标,众多学者开始致力于渔业生态模型的研究,希望借由虚拟模型模拟资源的发展态势,以此进行渔业管理和种群评估工作[5-6]。其中,由Shin等[7]提出的面向对象的海洋生态系统估计模型(object-oriented simulator of marine ecosystems exploitation,OSMOSE)已经应用于许多国家或地区的海洋生态系统研究中。如Travers-Trolet等[8]于2014年将高营养级模型OSMOSE与生物物理模型ROMS-N2P2Z2D2进行耦合,对东英吉利海峡海域水生态系统进行模拟研究,同时还探索了气候变化和捕捞压力等因素对生态系统及食物网的综合影响;
2020年,该作者用此模型估计了东英吉利海峡的最大可持续产量,结果与种群评估中得出的数值大体符合,其结论被用于捕捞建议[9]。在国内,Xing等[10]于2017年采用OSMOSE-JZB模型对胶州湾海域生态系统进行模拟研究;
2020年,Xing[11]又在此基础上用此模型评估了休渔期对胶州湾海域生态资源的保护作用,并量化了使用蒙特卡洛模拟方法对OSMOSE模型在评估鱼类群落、种群和个体状况时的影响[12]。
与其他模型相比,OSMOSE是端到端(end-to-end)的模型,它通过引入鱼类个体之间的捕食关系,得到了基于多物种和基于个体的鱼类模型,OSMOSE模型能够模拟从低营养级到高营养级物种的完整生态系统动态,适于大尺度和多物种的生态系统研究,因此,在支持EBFM方面具备较大优势。目前,OSMOSE网站(https://documentation. osmose-model.org)上已经公开了OSMOSE模型的源码程序及应用示例,研究者可根据研究需要利用OSMOSE模型或对其进行改进。
东海是中国重要的渔业活动聚集地,分布着舟山渔场等众多渔场[13],由于过度捕捞、污染和海洋开发等影响,该海域出现了生态结构变化、渔业资源衰竭等问题。如何转变目前的海洋渔业发展方式,调整管理、生产、资源保护间的关系,不仅是实现中国海洋生态文明建设、发展现代海洋经济的重要措施,对于争取和维护中国的海洋权益也具有重要意义[14]。另一方面,以往对东海生态系统的研究更多地聚焦于小范围的单一物种,缺乏对生态系统的整体性研究。
基于此,为了加强对东海渔业资源的管理,评估东海海洋生态系统的状况,制定合适的作业策略,恢复其生态系统的自然水平,本研究中建立了面向东海大尺度范围的OSMOSE模型(OSMOSE-ECS),对鲐Scomberjaponicus等东海海域19种常见的主要经济渔业物种的生活史进行模拟研究,分析了主要物种的生物量、平均营养级及平均体长等生活史参数的变动趋势,评估在建模数据有限的情况下,OSMOSE模型用于模拟大尺度东海海洋生态系统的动态过程。
本研究中选取东海26°~35°N、120°~130°E及其周边海域进行模型构建,图1中的网格即为本研究中OSMOSE-ECS模型的网格模块划分。为了更全面地描述东海海洋生态系统的组成,选取东海海域经济价值较大的种类,即常见的10种鱼类、7种甲壳类和2种头足类作为研究对象(表1),采用OSMOSE-ECS模型模拟各物种的生活史,对东海海洋生态系统的发展态势进行模拟和分析。
OSMOSE模型通过模拟生态系统间的食物网关系进行构建,即通过快速匹配低营养级(low trophic level,LTL)数据,使其与模型研究对象高营养级(high trophic level,HTL)生物耦合,从而完成构建。

本图基于自然资源部标准地图服务网站GS(2016)1562号标准地图为底图制作,底图边界无修改。The figure is based on the standard map GS(2016)1562 in the standard map service website of Ministry of Natural Resources of the People’s Republic of China, with no modifications of the boundaries in the standard map.图1 OSMOSE-ECS模型的研究区域及网格划分Fig.1 Study area and grid division of OSMOSE-ECS model

表1 研究对象Tab.1 Object species
2.1 LTL驱动设置
OSMOSE模型是以LTL作为驱动力运行的模型,LTL是高营养级种群的食物来源,通过直接输入netCDF文件的方式对LTL参数进行快速驱动配置。东海的初级生产力生物的种类和数量非常丰富,但东海大陆架的浮游生物和小型底栖动物等LTL的实际观测生物量数据却非常缺乏。为了更全面地反映整个研究区域的初级生产力情况,本研究中采用较具代表性的叶绿素a(Chl-a)浓度推算目标区域LTL的生物量[15]。
在特定海域中,可以根据Cadée提出的简化公式计算初级生产力[16]:
CChl-a=PS×E×D/2。
(1)
其中:CChl-a为初级生产力(mg/(m2·d));
PS为表层水中浮游植物的潜在生产力(mg/(m3·d));
E为真光层的深度(m);
D为每日白昼的时长(h/d)。其中,PS根据表层水中Chl-a的浓度计算,其计算公式为
PS=Ca×Q。
(2)
式中:Ca为表层Chl-a浓度(mg/m3);
Q为同化系数(mgC/(mgChl-a·h))。真光层E的深度取透明度的3倍,同化系数采用3.7[17]。
在OSMOSE-ECS模型的构建中,Chl-a浓度数据采用SeaWiFS(Sea-Viewing Wide Field-of-view Sensor)提供的2001—2010年海表Chl-a全球月平均气候态L3级产品,空间分辨率为9 km×9 km。该产品数据反演算法基于生物-光学统计算法模型[18],对于大洋一类水体的Chl-a浓度反演具有普遍可靠性。基于此,创建了研究区域的月度Chl-a浓度数据,其中时间步长设为0.5月。
2.2 HTL参数构建
OSMOSE模型基于个体模拟生态过程。从图2可见,模型本身包括6个模块,分别是与物种相关的系统模块、物种模块、年龄模块和超级个体模块,以及与物种空间分布相关的网格模块和单元模块[19]。其中,系统模块用于描述物种的群落结构,物种模块用于模拟物种的生活史,网格模块则包括了X×Y个单元模块,用于模拟物种的运动及位置变化。模型通过配置文件获取各研究物种的体长、死亡率等参数,在二维空间的X×Y个闭合网格中进行模拟。

标识为斜体的属性会随着时间的推移发生变化。
Attributes in italics can vary through time.图2 OSMOSE模型的结构示意图Fig.2 Structure of OSMOSE model
2.2.1 空间分布 模型中物种的分布和位置依靠二维空间的网格单元来定义,在OSMOSE-ECS模型中共设置了20×18个网格单元,即在研究区域内以0.5°的分辨率分别对经纬度进行划分(图1)。在实际情况中,物种在目标区域中的移动可能是不具备游泳能力时随洋流的变化产生的,也可能是具备游泳能力后因生活习性而产生的变化,在数据充足时,每一个物种在研究区域内的变化都应该是不同的。由于缺乏数据,在OSMOSE-ECS模型中,各个物种均设为能够以各自的运动能力在其分布的网格区域内任意游动,各目标物种的分布区域通过查阅相关文献获得[20]。
2.2.2 种群参数
1)觅食。OSMOSE-ECS模型中只考虑具备觅食能力的个体物种(可通过年龄或体长在参数中设定判断条件),在单位时间步长内,由于个体的移动都是随机的,受到其所在网格单元Cx0,y0及临近单元Cx,y的营养水平Vx0,y0的影响,满足一定条件后,就会产生觅食移动。其中,Vx0,y0可以通过下式计算:
Vx0,y0={Cx,y|(x,y)∈([x0-1,x0+1]×
[y0-1,y0+1])∩([0,X]×[0,Y])}。
(3)
当处于单元Cx0,y0的物种种群i0向单元Cx′,y′移动时,需要满足条件:
(4)
其中:nx,y为单元Cx,y中存在的种群数量;
Li为属于种群i的物种体长;
Bi,x,y为单元Cx,y中种群i的生物量[19]。
2)捕食。如果捕食者和被捕食者在相同的时间出现在相同的空间区域内,且捕食者与被捕食者的体长比例符合捕食标准的要求,那么在OSMOSE-ECS模型中就可能出现机会捕食行为。在OSMOSE-ECS模型中,可以通过设置配置文件中的捕食矩阵参数来表示一个物种被另一物种捕食的概率[21]。需要注意的是,虽然OSMOSE-ECS中个体在二维平面上运动,但有些物种因为所处的水层深度不同,所以即使在二维空间上是重叠的,但并不会相遇,也无法发生捕食,这种情况下可以将捕食矩阵中对应的捕食概率设置为较低的值,或者设置为0,以表示二者不会发生捕食行为。
在每个网格单元(x,y)中,单位时间步长t′内,物种或种群i能够获得的猎物生物量总量APi,x,y,t′可以通过下式计算:
(5)
(6)
其中:Bj,x,y,t′为单位时间步长t′内在网格单元(x,y)被捕获的物种的生物量;
ai,j为猎物j对捕食者种群i的被捕食系数;
sr为捕食者与猎物合适的体长比;
PBi,j,Δt为猎物j被捕食者种群i捕食的生物量;
ri为最大摄入比,本模型中设为3.5。
3)生长。在OSMOSE-ECS模型中,根据von Bertalanffy生长模型[22]计算鱼类体长的增长率。根据估计,鱼类的每日食物定量(即维持生物量既不增加也不减少的食物量)约为体质量的0.55%,也就是说,对个体而言,每年体质量增长1 g需要的食物定量为2.01 g,据此计算得到临界值。当进食量高于此临界值时,即可认为多出的食物配给是物种专门用于生长而摄取的,而当个体摄入的食物量低于物种的每日食物定量时,个体则可能因为饥饿而停止生长乃至死亡。
根据von Bertalanffy模型可以获得不同年龄段鱼类的体长,即:
ΔLs,a=L∞s(1-e-KΔt)e-K(a-t0s)。
(7)
其中:ΔLs,a表示a龄的物种s在Δt时间里增加的平均体长;
L∞s为物种s的渐近体长;
K为该物种的生长参数;
t0s为物种s的理论起始年龄。

(8)
其中:t′为时间步长;
cs为方程的参数。
4)繁殖。食物摄取量除与生长的变化相关外,在一定程度上与物种的繁殖过程有关,特别是每个种群在局部的时间和空间遇到的食物条件。因此,在直接模拟每个物种的补充量水平之外,OSMOSE-ECS还建立了产卵量与产卵生物量间的关系模型。幼鱼是成鱼大量的食物来源,对系统的动态影响较大,因而明确建立鱼类幼体阶段的模型非常重要;
模型中的繁殖过程也不能简单地用线性关系进行模拟,否则会缺少对种群补充量关系的约束,鱼卵和幼鱼的年存活率与捕食过程、年承载力水平相关,因此,对繁殖行为的模拟也很有意义。
单位时间步长t′内,物种s产生的鱼卵数量与产卵群体的生物量(SBs,t′)是成一定比例的,假设种群内的性别比为1∶1,则产卵数量(Ns,t′)为
Ns,t′=0.5×φs×SBs,t′×seasonality。
(9)
其中:φs为物种s的相对繁殖力;
seasonality为在一个模拟年内该时间步长中产卵的占比。
5)饥饿死亡率。当食物配给量过低,无法提供物种基本的生命维持需求而发生饥荒,或种群对环境变化和疾病的敏感性增加等情况发生时,物种就会面临饥饿死亡的问题。在OSMOSE-ECS模型中,种群i的饥饿死亡率Mξi通过下式计算:
Mξi=-Mξmax×ξi/ξcrit+Mξmax。
(10)
其中:Mξmax为最大饥饿死亡率;
ξi为种群的捕食效率;
ξcrit为产生饥饿死亡时的捕食效率阈值。
6)捕捞死亡率。OSMOSE-ECS模型中,捕捞死亡只会发生在大于等于捕捞尺寸的种群个体上,确定捕捞死亡发生的时间是估算捕捞死亡率的关键。在一定的时间步长内,每个种群的行动顺序和相互作用,会造成其环境中捕食猎物群的显著死亡,也可导致自身发生可变饥饿死亡。因此,可以认为捕食和饥饿死亡在同一时间步长内是连续的[23]。本模型中,没有通过设置休渔期、保护区等参数对捕捞死亡率进行限制,而是使得生态系统持续处于一定的捕捞强度。
除此之外,模型中也会考虑到幼体死亡(通过模型校准获得)和其他死亡(如疾病、被海鸟捕食等情况)等因素,这些因素可以作为未知量通过模型校准获得。
2.2.3 HTL参数化构建 OSMOSE-ECS模型中涉及的HTL共有19个物种,模型运行所需的参数以csv文件的格式保存并被模型读取,可以分为种群相关参数、随机游走参数、摄食参数和空间分布参数等。其中,各研究对象的大部分物种参数通过Fish Base[24]和相关文献获取,一些难以获取的参数进行了适当估计,并根据相关文献设置了物种间的捕食效率和被捕食的概率。OSMOSE-ECS模型所采用的主要HTL物种参数见表2。
2.3 模型的校准
由于OSMOSE模型的复杂结构,为了提高模型的收敛性,本研究中模型的校准以现有的观测数据作为基准与模型的输出结果进行对比,并通过进化算法对模型的未知参数进行估计,模型校准流程如图3所示。在观测数据中主要通过生物量、产量、个体数量和一些阈值进行校准,根据数据特点设置相应的校准类型,从而比较观测结果和模拟结果。未知参数根据与模型的相关性,按重要程度进行分段,先对重要参数进行估算,估算得到的参数参与下一分段共同估算[11-12]。
OSMOSE-ECS模型通过R语言的calibra包进行校准,在校准时根据观测数据的似然性对目标函数进行优化设置。

表2 OSMOSE-ECS模型中物种主要参数Tab.2 Specific parameters of species in OSMOSE-ECS model

图3 OSMOSE-ECS模型校准流程Fig.3 Calibration process of OSMOSE-ECS model
本研究中的OSMOSE-ECS模型根据3次模拟得到的平均结果作为输出,模拟时间设置为10年(2001—2010年),时间步长为0.5月,即共设240个步长。对于种群中的物种,采用随机死亡算法模拟死亡过程。模型最终输出各目标物种种群的营养级、生物量和平均体长等模拟结果,反映目标物种在10年内的种群参数变化情况。
3.1 平均营养级模拟结果
为了验证模型模拟平均营养级结果的可靠性,本研究中选取部分目标物种与现有文献资料中通过胃含物成分及氮同位素法测定的平均营养级数据进行对比(表3)。结果显示,模拟结果与氮同位素法的测定结果更为接近,但总体上模型的模拟结果与两种实测方式的结果相差不大,表明OSMOSE-ECS模型输出的营养级参数是可以应用于生态系统分析与研究的。

表3 部分研究对象平均营养级模拟结果与实测值的比较Tab.3 Comparison between simulation results and measured value of mean trophic level for some species
OSMOSE-ECS输出的各目标物种在模拟时间内平均营养级的变化趋势如图4所示。整体上看,目标生物的营养级集中在3.00~4.50,大部分目标物种的平均营养等级变化不大,受到模型中繁殖期设置等因素的影响,变化趋势呈现一定的周期性波动态势,且波动较为平稳,个别物种(如带鱼(SP3)、太平洋褶柔鱼(SP11)等)的波动相对较大,但仍在相对稳定的范围内。平均营养级最高的为龙头鱼(SP4)、海鳗(SP5)等肉食性鱼类,甲壳类生物(SP13~SP19)的平均营养级较低,营养级集中在3.00附近,比较符合实际情况。

图4 平均营养级模拟结果Fig.4 Simulation result of mean trophic level
图5为模拟时间内(2000—2010年)生物量加权后的平均营养级变化趋势。从图5可以看出:东海生态系统的平均营养级在模拟期内为3.10~3.90,平均营养级为3.41;
生物量平均营养级在2006—2007年达到最高值,说明在此期间高营养级生物在生态系统中占据优势地位,但随后很快发生了下降,较低营养级的物种又重新占据了主导,营养级变化也趋于平稳。这表明,目标海域的生物资源结构发生了较大变化。

图5 生物量平均营养级变化趋势Fig.5 Trend of mean trophic level by biomass
3.2 生物量模拟结果
OSMOSE-ECS模型生物量输出结果表示的是整个研究区域范围内该物种生物量的总和。本研究中选取了鲐、带鱼两个主要目标物种对模型模拟结果进行验证,通过与实际捕捞渔获量的变化趋势对比,验证OSMOSE-ECS模拟结果的可靠性。
图6为鲐模拟生物量与实际渔获量的变化趋势对比图,由于资料有限,本研究中截取了其中5年(2001年8月—2006年8月)模拟生物量与实际渔获量数据[27]进行对比。由于初始生物量设定的关系,模拟生物量与实际渔获量的单位数量级有所不同,为了便于分析,图中进行了放大处理。从图6可见,模拟生物量的变化趋势与实际渔获量的变化走向非常吻合。在各观测时间点计算实际渔获量与模拟生物量的皮尔逊相关系数约为0.47,可认为二者具有一定的相关关系。

图6 鲐模拟生物量与实际渔获量趋势对比Fig.6 A trend comparison between simulated biomass and real catching yield of chub mackerel Scomber japonicus
图7为带鱼的生物量模拟结果与实际渔获量[28]的变化趋势对比。从图7可见,带鱼的模拟生物量变化幅度较大,趋势规律性较弱,但与实际渔获量数据的走向也是相符的。图中选取的实际渔获量跨度为10年,可以看出,模拟值与实际值均先达到一个较高的峰值,之后逐渐走低,最后趋于平稳。同样地,在各观测时间点计算得到模拟生物量值与实际渔获量值的皮尔逊相关系数约为0.79,由于数据有限,观测时间点的数量和间隔不均匀,因此,在走势上虽未完全吻合,但依然能够反映出OSMOSE-ECS中带鱼生物量的模拟结果与实际产量具有相关性。

图7 带鱼模拟生物量与实际渔获量趋势对比Fig.7 A trend comparison between simulated biomass and real catching yield of hairtail Trichiurus japonicus
结合鲐与带鱼的模拟结果,可以认为OSMOSE-ECS输出的生物量值能够反映物种实际的生物量变化规律。图8给出了在模拟时间内模型中各目标物种模拟生物量的变化趋势,由于不同物种间的数据量差异较大,因此,选择了常用对数坐标进行表示。从图8可见,大部分目标物种的生物量在模拟开始后的20个月内就能够达到平稳波动,总生物量在较固定范围内振荡,并于模拟时间末期均趋于稳定。在选定的目标物种中,具有生态优势的几个物种(如海鳗(SP5)等)的生物量亦多于其他优势不明显的物种,符合其在东海生态系统中营养等级的位置,总体上符合生物种群发展的一般规律。

图8 各物种生物量模拟对数结果Fig.8 Simulation logarithmic results of each species biomass
3.3 平均体长模拟结果
由于多数物种的有效观测数据缺乏,为了验证结果的可靠性,本研究中将鲐(SP1)、带鱼(SP3)平均体长模拟结果的变化趋势与部分实际观测值[29-30]进行了比较。从图9可见,虽然观测时间点较少,无法完整体现两类物种实际体长的连续变化趋势,但无论是模拟值还是实际观测值,鲐、带鱼的平均体长均发生了显著下降,数值范围也相差不大。
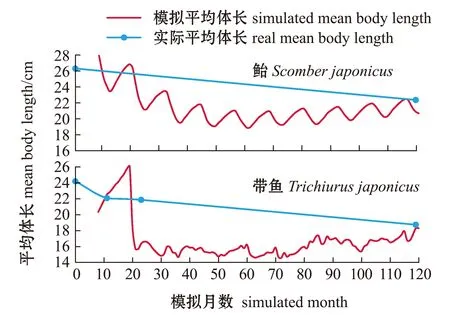
图9 鲐、带鱼的平均体长趋势模拟结果与实际数据趋势对比Fig.9 A trend comparison between simulated and real mean body length of chub mackerel Scomber japonicus and hairtail Trichiurus japonicus
图10为OSMOSE-ECS模型输出的各鱼类目标物种(SP1~SP10)的平均体长变化情况。从图10可见,大部分经济鱼类的体长在模拟时间范围内均呈现波动中缓慢下降的趋势,体长结构也相对偏小。在模拟结果中,仅营养级较高的蓝点马鲛(SP10)和海鳗(SP5)的平均体长有较大的升高趋势,这可能是模型参数设置的随机性(如捕食关系)产生的,但其体长仍在合理范围之内。

图10 主要经济鱼类平均体长模拟结果Fig.10 Simulation results of mean body length for main commercial fish
4.1 模型的选择与建立
OSMOSE模型作为端到端的模型,对观测数据的依赖较低,模型能够建立多物种间的关系,从而更完整地对海洋生态系统进行模拟。本研究中,构建了面向东海渔业生态系统的OSMOSE-ECS模型,与实际观测数据相比,OSMOSE-ECS模型的大部分模拟输出结果与实际观测数据在趋势上基本一致,说明利用OSMOSE模型对东海海洋渔业资源管理和生态评估提供支持是可行的。
长久以来,国内对于东海海域生态系统的研究大多面向个体,较少考虑生态系统综合因素对物种的影响。例如,采用MaxEnt模型对中国近海的石首鱼科鱼类进行研究[31],对中国近海带鱼科空间分布、种群结构与营养生态进行研究[32],以及对近海鲐[33]、龙头鱼[34]、葛氏长臂虾[35]等物种的研究等,这些研究较少考虑生态系统内部物种间复杂的关系机制。而本研究中的OSMOSE-ECS模型的优势,在于能够建立多物种之间的关系,实现对物种间捕食、竞争等关系的模拟。另一方面,现有的一些海洋生态系统模型研究多面向小尺度范围的生态系统,如采用OSMOSE模型,研究南非的南本格拉湾[7-9]、中国胶州湾[10-12]和秘鲁的上升流海域[15]等,尺度范围均不大。而本研究中将OSMOSE模型应用于东海这种较大尺度的海洋生态系统中进行模拟,并验证了该模型广泛的适应性。
由于实际观测参数获取有限,在本次模拟中,虽然采用叶绿素a浓度推算了初级生产力情况,但与真实情况还存在一定差异。此外,在模型构建的过程中也忽略了一些差异因素,如物种洄游产生的空间分布变化及伏季休渔等因素对生态系统的影响;
本研究中也存在模拟时间较短的问题,虽然一定程度上已经反映出了各目标物种的发展态势,但若延长模拟时间,各物种的发展趋势可能更加稳定,生态系统也能够达到更平衡的状态,数据的可靠性也会大大增强。在后续研究中,笔者将考虑改善LTL的表示方式,延长模型模拟的时间,对模型进行更全面的校准和敏感性分析,同时考虑引入休渔期、保护区等影响因子,以期能够更真实、准确地描述东海海洋生态系统的结构与状态。
4.2 OSMOSE-ECS模型对东海生态系统模拟的可行性
OSMOSE模型通过对个体之间的捕食关系进行模拟,可得到基于多物种的生态系统模型,能够模拟从低营养级到高营养级物种的完整生态系统动态,适于大尺度和多物种的生态系统研究。与现有研究相比,本研究中,OSMOSE-ECS模型模拟得到的2001年生物量平均营养级(3.53)与观测数据中2000—2001年间东海90%的生物量集中营养级(3.69)[26]非常接近,但模型模拟时间末期生物量平均营养级已经降低为3.16。从单一的物种来看平均营养级的波动并不大,但整体上东海生态系统中高营养级生物个体减少,包括各种鱼类(SP1~SP10)和头足类(SP11~SP12),整体平均营养级的下降可能源于食物的缺乏,也与人类的渔业捕捞活动密切相关。实际捕捞作业中,即使在营养级相对较低的甲壳类(SP13~SP19)中,捕捞量的增加及捕捞工具的改进都会使得生态系统中物种的优势度发生较大变化,进而破坏渔业资源[36]。
OSMOSE-ECS模型对生物量的模拟结果与实际渔获量观测值的走向较为一致,模拟结果中生物量的整体情况相对比较平稳,大部分鱼类生物量的波动幅度较大,呈现出明显的周期性;
头足类生命周期较短,种群的世代结构不连续,资源更新频率快[37],模拟结果中物种之间差异较大;
甲壳类各物种个体体质量较小,因此,虽然个体数量更多,整体生物量水平却较低,波动也不明显。模型输出的总生物量模拟值基本在一定范围内波动,并于模拟时间的末期均趋于稳定。本研究中的模拟时长较短,若在后续的研究中延长模拟时间,生物量数据最后应当能够更加平稳。
与平均营养级、生物量等模拟结果类似,OSMOSE-ECS模型对鱼类平均体长的模拟结果和实际观测数据均反映出除营养级相对较高的物种之外,各鱼种平均体长在不断下降,模拟数据的体长变化差距与实际观测数据也接近。同样地,从模拟结果可以看出,大部分目标物种的平均体长都发生了显著的下降,结合平均营养级和生物量的变化趋势,可以认为,在不存在休渔期或保护区、整个生态系统长期处于高强度捕捞作业的背景下,成年的、较大体型的生物个体则越来越少。以鲐(SP1)和带鱼(SP3)为例,二者的生物量和平均体长都有明显的下降,如果继续保持这样的捕捞强度,资源的开发潜力会进一步缩减。
从本文模拟结果总体来看,在模拟期间内东海生态系统种群结构发生了较大变化,单一物种平均营养变化并不大,但平均营养级下降,尤其是东海生态系统中高营养级生物个体比例正在减少;
对于大部分鱼类来说,单一生物量呈现出明显的周期性的波动,但总生物量平均水平变化比较平稳;
大部分经济鱼类的体长都呈现出在波动中缓慢下降的趋势,整体体长结构也相对偏小。这表明,东海生态系统的种群结构正在向低营养级的方向偏移。
综上,东海生态系统的营养等级相对偏低,这种不设休渔期或保护区的高强度捕捞作业,更加不利于生态系统的稳定。如果捕捞压力过大,很容易使鱼类种群的遗传结构发生变化,产生性早熟的趋势,以适应人为捕捞带来的种群延续压力,因此,体长较小、生长快、繁殖周期短的个体会占据主流[32]。若不改变现有捕捞方式,生物资源必然会继续下降,整个东海海洋生态系统也会因此受到影响发生较大的变化,届时渔业作业也很难获取到经济价值较高的个体[38]。
4.3 对东海生态系统渔业管理的建议
本研究中建立的模型在模拟过程中探讨了高强度捕捞作业下东海生态系统可能的发展情况。模拟结果显示,东海的生态系统正在衰退,可能的原因是模型中采用了东海区实际的捕捞死亡率,捕捞压力使得东海海洋生态系统的物种整体趋向小型化、低龄化,生态系统的稳定性大为降低,这也暴露出目前东海渔业管理上存在诸多问题。如果继续保持当前的捕捞强度,势必会使本已占据优势地位的经济鱼种发生衰退。东海渔业生态系统在承受过度的捕捞压力下会面临失衡风险,亟待调节。
基于此,作者认为,有必要对东海区的渔业政策进行适当调整。一方面,对东海生态系统的营养级水平进行长期、系统地监测和研究,同时,调整捕捞策略,根据不同物种的生物量实施专门的配额捕捞制度,根据不同物种的繁衍周期针对性地调整休渔期,或通过降低捕捞强度、改变开捕体长和设立保护区等措施,促进生态系统的恢复,保护各物种种群的生长和繁殖。另一方面,在立法上也需完善相关政策制度,促进利益相关者协力合作,以使东海生态系统健康、良性地发展,保障渔业资源能被长期、稳定、可持续地获取和利用。
1)OSMOSE-ECS模型对于尺度较大、物种关系较复杂的东海海洋生态系统能实现较好的模拟,可有效模拟各目标物种的平均营养级、生物量和平均体长等数据变化趋势。
2)根据模型模拟结果,在未对捕捞作业进行限制的条件下,模拟时间段内各目标物种的平均营养级变化相对平稳,但东海生态系统总生物量平均营养级呈下降趋势;
各目标物种的生物量呈现明显的周期性波动;
大部分鱼类目标物种的平均体长降低。该模型反映出模拟状态下东海生态系统中高营养级生物正在减少,物种个体趋向于小型化、低龄化,生态系统正在衰退。